

FAMILIENLISTE / Pterosauria

Kamp, 1834
Archosauria Pterosauria
Die am frühesten datierten Fossilfunde von Flugsauriern (Pterosauria, für "geflügelte Echsen") stammen aus der Trias vor ungefähr 231 bis 225 Millionen Jahren. Am Ende der Kreidezeit vor etwa 66 Millionen Jahren, verschwanden sie zusammen mit den meisten Dinosaurierarten. Insgesamt existierte diese Gruppe also beinahe 200 Millionen Jahre erfolgreich auf unserem Planeten. Zu der bekanntesten Art zählt wohl der Pteranodon. Daneben gab es aber noch eine Vielzahl von anderen Arten, die auf allen Kontinenten verbreitet waren. Die meisten der gefundenen Arten lebten in Meeresnähe, wahrscheinlich ernährten sie sich daher von Fisch.
Eine der wichtigen Fundstätten für Flugsaurierfossilien ist der Solnhofener Plattenkalk in Bayern. Bereits im frühen 18. Jahrhundert wurden hier die ersten Überreste gefunden, aber erst 1801 erkannte George Cuvier, dass es sich um flugfähige Reptilien handelte und nicht etwa um Fledermäuse. Durch die Fossilienfunde offenbart sich uns aber nur ein kleiner Ausschnitt der Artenvielfalt, da alle Versteinerungen praktisch Zufälle sind. Dazu kommt, dass Fossilien nur in Sedimentgestein entstehen können, welches im Bereich früherer Meere gefunden wird. Flugsaurier existierten ähnlich lange wie die Vögel heute und wurden auch weltweit nachgewiesen. Es ist daher wahrscheinlich, dass Flugsaurier auch im Landesinneren vorkamen, wie die Vögel. Das größte bis jetzt gefundene Fossil ist der Quetzalcoatlus mit einer Flügelspannweite bis zu 11 m, während einige andere Gattungen nicht größer als Sperlinge waren. Die ältesten Flugsaurier wurden in Italien (Eudimorphodon) und in Brasilien (Faxinalipterus) gefunden.

Zhejiangopterus / © Raul Lunia
Eines der markantesten Merkmale eines Flugsauriers sind die Proportionen des Skeletts: die Glieder und der Kopf waren sehr groß, während der Oberkörper im Vergleich dazu recht klein war. Dieser Trend war am extremsten bei einigen der großen Pterodactyloiden aus der Kreidezeit, aber auch frühe Flugsaurier haben Proportionen, die wir (als Säugetiere) sehr seltsam finden. Innerhalb der oft enormen Schädel der Flugsaurier sind mehrere wichtige Öffnungen zu vorhanden. Eines der auffälligsten waren die antorbitalen Fenster im Schädel vor der Augenhöhle. Bei einigen der pterodactyloiden Arten reicht diese Öffnung bis in die Nasenöffnungen, um eine rundes, nasoantorbitales Fenster zu bilden. Diese gemeinsame Öffnung im Schädel war bei vielen Flugsauriern der Kreidezeit, vor allem bei verschiedenen azhdarchoiden Arten enorm. Eine solche Ausdehnung von Öffnungen und Durchgängen im Skelett unterstützte die Umverteilung von Gewicht - die Arten mit großen nasoantorbitalen Fenstern neigten auch dazu, ein riesiges Schädelvolumen zu entwickeln. Die mögliche Erhöhung der Skelettmasse aus der Erweiterung des Schädels wurde so teilweise durch den Ausbau der Öffnungen im Schädel ausgeglichen. Die hochfeste Struktur ihrer Knochen ermöglichte es Pterosauriern, sowohl sehr stark als auch sehr leicht zu sein.
Der primäre Flügel wird als Brachiopatagium (Armflügel) bezeichnet. Die Flügel der Pterosaurier wurden durch Hautmembranen und anderen Geweben gebildet. Die Membranen erstreckten sich vom extrem langen vierten Finger entlang des Armes bis zu den Körperseiten. Die Flügelmembranen der Flugsaurier mit ihren sehr komplexen und dynamischen Strukturen waren zu einem aktiven Flugstil sehr gut geeignet. Die äußeren Flügel (von der Spitze bis zum Ellenbogen) wurden von eng beieinander liegenden Fasern (Aktinofibrillen) verstärkt. Diese Fasern bestanden aus drei unterschiedlichen Schichten, die sich im Flügel kreuzweise überlagerten. Die eigentliche Funktion der Fasern ist unbekannt, ebenfalls wie das Material, aus dem sie bestanden. Je nach ihrer genauen Zusammensetzung (Keratin, Muskeln, elastische Strukturen etc.), könnten sie zur Stabilisierung des äußeren Flügelteils gedient haben. Die Flügelmembranen selbst bestanden aus einer dünnen Schicht aus Muskeln und Bindegewebe, die durch ein komplexes System von Blutgefäßen durchzogen waren. Hohlräume in den Flügelknochen bei den größeren Arten besaßen luftgefüllte Bereiche, die das Gewicht erheblich reduzierten.

Rhamphorhynchus / © Raul Lunia
Die Rekonstruktion aus fossilen Knochen ist keine leichte Aufgabe. Und um zu erkennen, wie ein Tier atmete, erst recht nicht. Aber es gibt bei den Flugsauriern viele Hinweise darüber. Einige der Knochen waren hohl, von vielen (wenn nicht den meisten) Arten, weiß man, dass an die Atemwege Luftsäcke angeschlossen waren. Die besondere Verzahnung der Rippen und das Brustbein legen nahe, dass sie die Brust in ähnlicher Weise bewegten wie die Vögel. Zusammengenommen zeigen diese Faktoren, dass Flugsaurier wahrscheinlich ein sehr effizientes Lungensystem ähnlich dem der Vögel besaßen. Dies würde bedeuten, dass den Flugsauriern wahrscheinlich die Membran fehlte (unser Hauptatemmuskel). Sie verwendeten wahrscheinlich eine komplexe pulsierende Bewegung des Brustbeins und der Brustwand, um ihre Atmung zu unterstützen. Es ist durchaus möglich, dass die größten Pterosaurier eine große Menge von dem, was als "anaerobe Leistung" bezeichnet wird, verwendet haben. Das heißt, viel von ihrer Muskelmasse bestand aus Hochleistungsfasern, die nicht viel Sauerstoff benötigten um Kraft zu erzeugen, aber sehr schnell ermüdeten.

Pterosaurier-Portraits / © Maija Karala
Die meisten Flugsaurier-Fossilien enthalten zumeist keine Abdrücke der Hautoberfläche. Von einigen Exemplaren, ist bekannt, dass sie eine teilweise haarähnliche Körperbedeckung besaßen, die ihnen ein pelziges Aussehen verlieh. Einige Exemplare wie Jeholopterus scheinen mit so einem "Fell" ausgestattet gewesen zu sein. Was die Funktion und Wirkung dieser Hautbedeckungen bewirkte, ist schwer abzuschätzen. Es ist aber wahrscheinlich, dass sie zum Teil der Isolierung dienten. Das wäre ein zusätzlicher Hinweise darauf, dass Flugsaurier schnelle Stoffwechselraten besaßen, also Warmblüter waren.
Die Flugsaurier werden als Diapsiden in die Klasse der Reptilien gestellt. Innerhalb der Klasse sind sie den Archosauriern zugeordnet, die auch Dinosaurier und Krokodile beinhaltet. Die Pterosaurier werden in die Unterordnungen Rhamphorhynchoidea (Langschwanzflugsaurier) und die Pterodactyloidea (Kurzschwanzflugsaurier) eingeteilt. Die ersteren waren dabei die früheren Formen, sie lebten im Trias und Jura. Die Kurzschwanzflugsaurier lebten im Jura und spalteten sich vermutlich von den frühen Formen ab. Es gibt aber keine Fossilfunde von Vorläufern.
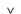 Pterosauria
Pterosauria
. Allkaruen
. Arcticodactylus
. Austriadraco
. Comodactylus
. Daohugoupterus
. Peteinosaurus
. Pterorhynchus
. Seazzadactylus
. Sordes
- Monofenestrata (Lü, Unwin, Jin, Liu, Ji, 2010)
-. Douzhanopterus
")
© Uwe Jelting")
© Mark Witton
© Raul Lunia
")
Rhamphorhynchus
© Claessens et al.")
Pterosauriers
© John Conway
Pterosauriers
© Naish et al.
Weitere Informationen
A Jurassic pterosaur from Patagonia and the origin of the pterodactyloid neurocranium / Laura Codorniú, Ariana Paulina Carabajal, Diego Pol, David Unwin, Oliver W.M. Rauhut, 2016 / PeerJ 4:e2311 https://doi.org/10.7717/peerj.2311 /
/ Laura Codorniú, Ariana Paulina Carabajal, Diego Pol, David Unwin, Oliver W.M. Rauhut, 2016 / PeerJ 4:e2311 https://doi.org/10.7717/peerj.2311 / PDF; Ergänzendes Material PDF
PDF; Ergänzendes Material PDF
Air Space Proportion in Pterosaur Limb Bones Using Computed Tomography and Its Implications for Previous Estimates of Pneumaticity / Elizabeth G. Martin, Colin Palmer, 2014 / PLoS ONE 9(5): e97159. doi:10.1371/journal.pone.0097159 /PDF
An analysis of pterosaurian biogeography: implications for the evolutionary history and fossil record quality of the first flying vertebrates / Paul Upchurch, Brian Andres, Richard J. Butler, Paul M. Barrett, 2015 / Historical Biology, Volume 27, Issue 6, Pages 697-717PDF
A new pterosaur tracksite from the Lower Cretaceous of Wuerho, Junggar Basin, China: inferring the first putative pterosaur trackmaker / Yang Li, Xiaolin Wang, Shunxing Jiang, 2021 / PeerJ 9:e11361 https://doi.org/10.7717/peerj.11361PDF
Allometric wing growth links parental care to pterosaur giantism / Zixiao Yang, Baoyu Jiang, Michael J. Benton, Xing Xu, Maria E. McNamara, David W. E. Hone, 2023 / Proc. R. Soc. B. 2902023110220231102. https://doi.org/10.1098/rspb.2023.1102 /PDF
Arthrological reconstructions of the pterosaur neck and their implications for the cervical position at rest / Richard Buchmann, Taissa Rodrigues, 2024 / PeerJ 12:e16884 https://doi.org/10.7717/peerj.16884 /PDF
Comments on Triassic pterosaurs with discussion about ontogeny and description of new taxa/ Alexander W. A. Kellner, 2015 / An. Acad. Bras. Ciênc. Vol. 87 no.2 Rio de Janeiro Apr./June 2015 /PDF
Comparative analysis of the vertebral pneumatization in pterosaurs (Reptilia: Pterosauria) and extant birds (Avialae: Neornithes)/ Richard Buchmann, Leonardo dos Santos Avilla, Taissa Rodrigues, 2019 / PLoS ONE 14(10): e0224165. https://doi.org/10.1371/journal.pone.0224165 /PDF
Early Cretaceous pterosaur tracks from a “buried” dinosaur tracksite in Shandong Province, China / Li-Da Xing, Jerald D. Harris, Gerard D. Gierlinski, Murray K. Gingras, Julien D. Divay, Yong-Gang Tang, Philip J. Currie, 2012 / Palaeoworld 21 (2012) 50–58
/ Li-Da Xing, Jerald D. Harris, Gerard D. Gierlinski, Murray K. Gingras, Julien D. Divay, Yong-Gang Tang, Philip J. Currie, 2012 / Palaeoworld 21 (2012) 50–58
Evolution of the pterosaur pelvis / Elaine S. Hyder, Mark P. Witton, David M. Martill, 2014 / Acta Palaeontologica Polonica 59 (1), 2014: 109-124 doi: http://dx.doi.org/10.4202/app.2011.1109 /PDF
Filter feeding in Late Jurassic pterosaurs supported by coprolite contents/ Martin Qvarnström, Erik Elgh, Krzysztof Owocki, Per E. Ahlberg, Grzegorz Niedźwiedzki, 2019 / PeerJ 7:e7375 https://doi.org/10.7717/peerj.7375 / PDF
Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary/ Nicholas R. Longrich, David M. Martill, Brian Andres, 2018 / PLoS Biol 16(3): e2001663. https://doi.org/10.1371/journal.pbio.2001663 /PDF
Morphometric assessment of pterosaur jaw disparity/ Charlie A. Navarro, Elizabeth Martin-Silverstone, Thomas L. Stubbs, 2018 /
Royal Society Open Science, http://doi.org/10.1098/rsos.172130 /PDF
New material of the non-pterodactyloid pterosaur Changchengopterus pani Lü, 2009 from the Late Jurassic Tiaojishan Formation of western Liaoning / Chang-Fu Zhou, Rainer R. Schoch, 2011 / Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen Band 260 Heft 3 (2011), p. 265 - 275
New postcranial remains from the Lealt Shale Formation of the Isle of Skye, Scotland, showcase hidden pterosaur diversity in the Middle Jurassic / Natalia Jagielska, Thomas J. Challands, Michael O'Sullivan, Dugald A. Ross, Nicholas C. Fraser, Mark Wilkinson, Stephen L. Brusatte, 2023 / Scottish Journal of Geology (2023) 59 (1-2): sjg2023-001. https://doi.org/10.1144/sjg2023-001 /PDF
On the supposed pterosaurian nature of Faxinalipterus minima Bonaparte et al. (2010) from the Upper Triassic of Rio Grande do Sul, Brazil./ Marina B Soares, Fabio Marco Dalla Vecchia, Cesar Leandro Schultz, Alexander W. A. Kellner, 2013 / Conference: Rio Ptero 2013 - International Symposium on pterosaurs, At Rio de Janeiro
Potential For Intracranial Movements in Pterosaurs / Edina Prondvai, Attila Ösi, 2011 / Anatomical Record, 294: 813–830. doi:10.1002/ar.21376
Powered flight in hatchling pterosaurs: evidence from wing form and bone strength / Darren Naish, Mark P. Witton, Elizabeth Martin-Silverstone, 2021 / Scientific Reports, Volume 11, Article number: 13130 (2021)PDF
Pterosaur remains from the Kayenta Formation (?early Jurassic) of Arizona / Kevin Padian, / Palaeontology 27(2), pp. 407-413
Pterosauria of the Great Oolite Group (Bathonian, Middle Jurassic) of Oxfordshire and Gloucestershire, England/
Michael O’Sullivan, David M. Martill, 2018 / Acta Palaeontologica Polonica 63 (4), 2018: 617-644 doi:https://doi.org/10.4202/app.00490.2018 /PDF
Pterosaur teeth from the Lower Cretaceous (Valanginian) Cliff End Bone Bed, Wadhurst Clay Formation, Wealden Supergroup of southern England, and their possible affinities / Steven C. Sweetman / Cretaceous Research 151 (2023) 105622 /PDF
Respiratory Evolution Facilitated the Origin of Pterosaur Flight and Aerial Gigantism/ Leon P. A. M. Claessens, Patrick M. O'Connor, David M. Unwin, 2009 / PLoS ONE 4(2): e4497. doi:10.1371/journal.pone.0004497 /PDF
Seazzadactylus venieri gen. et sp. nov., a new pterosaur (Diapsida: Pterosauria) from the Upper Triassic (Norian) of northeastern Italy / Fabio Marco Dalla Vecchia, 2019 / PeerJ 7:e7363 https://doi.org/10.7717/peerj.7363 /PDF
Testing pterosaur ingroup relationships through broader sampling of avemetatarsalian taxa and characters and a range of phylogenetic analysis techniques / Matthew G. Baron, 2020 / PeerJ 8:e9604 https://doi.org/10.7717/peerj.9604PDF
The extent of the pterosaur flight membrane/ Ross A. Elgin, David W.E. Hone, Eberhard Frey, 2011 / Acta Palaeontologica Polonica 56 (1), 2011: 99-111 doi: http://dx.doi.org/10.4202/app.2009.0145 /PDF
Were early pterosaurs inept terrestrial locomotors?/ Mark Witton, 2015 / PeerJ 3:e1018; DOI 10.7717/peerj.1018 /PDF
- - - - -
Bildlizenzen
Pterodactylus © Uwe Jelting:
") Creative Commons 4.0 International (CC BY 4.0)
Creative Commons 4.0 International (CC BY 4.0)
Quetzalcoatlus & Giraffe © Mark Witton:
") Creative Commons NonCommercial-ShareAlike 2.0 Generic (CC BY-NC-SA 2.0)
Creative Commons NonCommercial-ShareAlike 2.0 Generic (CC BY-NC-SA 2.0)
Atem-Kinematik des Rhamphorhynchus © Claessens et al.
Creative Commons 4.0 International (CC BY 4.0)
Struktur eines Pterosauriers © John Conway:
") Creative Commons NonCommercial-NoDerivs 3.0 Unported (CC BY-NC-ND 3.0)
Creative Commons NonCommercial-NoDerivs 3.0 Unported (CC BY-NC-ND 3.0)
Entwicklung eines Pterosauriers © Naish et al.:
Creative Commons 4.0 International (CC BY 4.0)
- - - - -
Grafiken und Illustrationen von Raul Lunia
Grafiken und Illustrationen von Maija Karala